For det utrente øje ser planerne muligvis ud til at vokse temmelig impulsivt, og de løber tilfældigt ud og skaber et stort grønt virvar. Dog kigge nærmere på, og du vil opdage, at et par nysgerrig regelmæssige mønstre dukker op overalt i den naturlige verden, fra den afbalancerede symmetri af bambusskud til de betagende spiraler af sukkulenter.
Faktisk er disse mønstre konstante nok til, at kold, hård matematik kan forudsige organisk vækst temmelig godt. En antagelse, der har været central for studiet af phyllotaxis eller bladmønstre, er, at blade beskytter deres personlige rum. Baseret på ideen om, at allerede eksisterende blade har en hæmmende indflydelse på nye, hvilket giver et signal for at forhindre andre i at vokse i nærheden, har forskere skabt modeller, der med succes kan genskabe mange af naturens fælles design. Den stadigt fascinerende Fibonacci-sekvens viser sig for eksempel i alt fra solsikkefrøarrangementer til nautilusskaller til fyrretræer. Den nuværende konsensus er, at bevægelserne af væksthormonet auxin og de proteiner, der transporterer det gennem en plante, er ansvarlige for sådanne mønstre.
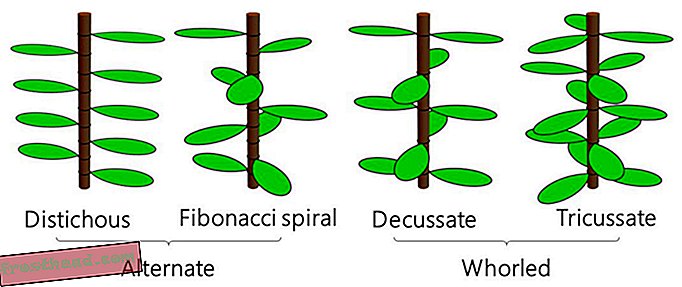 Bladarrangement med et blad pr. Knude kaldes alternativ phyllotaxis, mens arrangement med to eller flere blade pr. Knude kaldes whorled phyllotaxis. Almindelige alternative typer er distichous phyllotaxis (bambus) og Fibonacci spiralphyllotaxis (den sukkulente spiral aloe), og almindelige whorled typer er decussate phyllotaxis (basilikum eller mynte) og tricussate phyllotaxis ( Nerium oleander, undertiden kendt som dogbane). (Takaaki Yonekura under CC-BY-ND)
Bladarrangement med et blad pr. Knude kaldes alternativ phyllotaxis, mens arrangement med to eller flere blade pr. Knude kaldes whorled phyllotaxis. Almindelige alternative typer er distichous phyllotaxis (bambus) og Fibonacci spiralphyllotaxis (den sukkulente spiral aloe), og almindelige whorled typer er decussate phyllotaxis (basilikum eller mynte) og tricussate phyllotaxis ( Nerium oleander, undertiden kendt som dogbane). (Takaaki Yonekura under CC-BY-ND) Visse bladarrangementer fortsætter imidlertid med at stubbe populære modeller for plantevækst, herunder Douady- og Couder-ligningerne (kendt som DC1 og DC2), der har domineret siden 1990'erne. Et team ledet af University of Tokyo-forskere, der studerede en busk kendt som Orixa japonica, fandt, at tidligere ligninger ikke kunne genskabe plantens usædvanlige struktur, så de besluttede at genoverveje selve modellen. Deres opdaterede model, der er beskrevet i en ny undersøgelse i PLOS Computational Biology, gengiver ikke kun det engang svækkende mønster, men det kan også beskrive andre, mere almindelige arrangementer bedre end tidligere ligninger, siger forfatterne.
"I de fleste planter har phyllotactic mønstre symmetri - spiralsymmetri eller radial symmetri, " siger University of Tokyo plantefysiolog Munetaka Sugiyama, seniorforfatter til den nye undersøgelse. ”Men i denne specielle plante, Orixa japonica, er det phyllotaktiske mønster ikke symmetrisk, hvilket er meget interessant. For mere end 10 år siden kom en idé til mig om, at nogle ændringer i den hæmmende kraft af hvert blad Primordium kan forklare dette ejendommelige mønster. ”
Botanikere bruger divergensvinkler eller vinkler mellem på hinanden følgende blade til at definere en plantes phyllotaxis. Mens de fleste bladarrangementmønstre holder en konstant divergensvinkel, vokser O. japonica- busken, der er hjemmehørende i Japan og andre dele af Østasien, blade i en skiftevis række på fire gentagne vinkler: 180 grader, 90 grader, 180 grader igen, derefter 270 grader.
 En Orixa japonica busk med de forskellige divergensvinkler på bladene synlige. (Qwert1234 via Wikicommons under CC BY-SA 4.0)
En Orixa japonica busk med de forskellige divergensvinkler på bladene synlige. (Qwert1234 via Wikicommons under CC BY-SA 4.0) Dette mønster, som forskerne kaldte ”orixat” phyllotaxis, er ikke kun en engangsforstyrrelse, da planter fra andre taxaer (som ”rød-varm poker” -blomsten Kniphofia uvaria eller crepe myrtle Lagerstroemia indica ) skifter deres blade i det samme kompliceret rækkefølge. Fordi bladarrangementet dukker op forskellige steder på det evolutionære træ, konkluderede forfatterne, at ligheden kom fra en fælles mekanisme, der berettigede til yderligere undersøgelse.
Efter test af Douady- og Couder-ligningerne med forskellige parametre kunne forfatterne fremstille mønstre, der var tæt på det vekslende orixatarrangement, men ingen af de simulerede planter stemte perfekt sammen med O. japonica- prøverne, de dissekerede og studerede. Så teamet byggede en ny model ved at tilføje en anden variabel til Douady og Couder-ligningerne: bladalder. Tidligere modeller antog, at blade 's hæmmende kraft forblev den samme over tid, men denne konstante var "ikke naturlig ud fra biologiens synspunkt, " siger Sugiyama. I stedet tillader Sugiyama's team muligheden for, at styrken af disse “holde sig væk” -signaler ændrede sig over tid.
De resulterende modeller - som teamet omtaler som udvidede Douady- og Couder-modeller, EDC1 og EDC2 - lykkedes ved at genoprette, gennem edb-vækst, de intrikate bladarrangementer af O. japonica . Ud over dette resultat producerede de udvidede ligninger også alle de andre almindelige løvmønstre og forudsagde de naturlige frekvenser af disse sorter mere nøjagtigt end tidligere modeller. Især i tilfælde af spiralformede planter forudsagde den nye EDC2-model Fibonacci-spiralen “superdominans” sammenlignet med andre arrangementer, mens tidligere modeller ikke kunne forklare, hvorfor denne særlige form ser ud til at vises overalt i naturen.
”Vores model, EDC2, kan generere orixatmønstre ud over alle hovedtyper af phyllotaxis. Dette er helt klart en fordel i forhold til den tidligere model, ”siger Sugiyama. "EDC2 passer også bedre til den naturlige forekomst af forskellige mønstre."
 Blade på en Orixa japonica gren (øverst til venstre) og et skematisk diagram over orixat phyllotaxis (højre). Det orixate mønster viser en særegen fircyklusændring af vinklen mellem blade. Et scanningselektronmikroskopbillede (midt og nederst til venstre) viser vinterknoppen af O. japonica, hvor blade først begynder at vokse. Primordiale blade er mærket i rækkefølge med det ældste blad som P8 og det yngste blad som P1. Etiketten O markerer skyderens spids. (Takaaki Yonekura / Akitoshi Iwamoto / Munetaka Sugiyama under CC-BY)
Blade på en Orixa japonica gren (øverst til venstre) og et skematisk diagram over orixat phyllotaxis (højre). Det orixate mønster viser en særegen fircyklusændring af vinklen mellem blade. Et scanningselektronmikroskopbillede (midt og nederst til venstre) viser vinterknoppen af O. japonica, hvor blade først begynder at vokse. Primordiale blade er mærket i rækkefølge med det ældste blad som P8 og det yngste blad som P1. Etiketten O markerer skyderens spids. (Takaaki Yonekura / Akitoshi Iwamoto / Munetaka Sugiyama under CC-BY) Forfatterne kan endnu ikke konkludere, hvad der præcist får bladalderen til at påvirke disse vækstmønstre, selvom Sugiyama spekulerer i, at det kan have at gøre med ændringer i auxintransportsystemet i løbet af en plantes udvikling.
Sådanne mysterier kunne løses ved ”push and pull” mellem computermodeller og laboratorieeksperimenter, siger Ciera Martinez, en computerbiolog, der ikke var involveret i undersøgelsen. Forfatterenes model giver et spændende skridt hen imod en bedre forståelse af phyllotaxis og giver plads til, at andre botanikere kan udfylde hullerne med plantesektion og analyse.
”Med modeller, selvom vi måske ikke kender den nøjagtige mekanisme endnu, får vi i det mindste kraftige ledetråde til, hvad vi skal kigge efter, ” siger Martinez i en e-mail. ”Nu skal vi bare se nærmere på molekylmekanismerne i virkelige planter for at prøve at finde ud af, hvad modellen forudsiger.”
 Et ovenfra og nedbillede af bladarrangementmønstre i "orixat" phyllotaxis som nye blade (røde halvcirkel) dannes fra skyde-spidsen (den sorte sorte cirkel) og vokser udad (Takaaki Yonekura under CC-BY-ND)
Et ovenfra og nedbillede af bladarrangementmønstre i "orixat" phyllotaxis som nye blade (røde halvcirkel) dannes fra skyde-spidsen (den sorte sorte cirkel) og vokser udad (Takaaki Yonekura under CC-BY-ND) Sugiyamas team arbejder for at forfine deres model endnu mere og få den til at generere alle kendte phyllotactic mønstre. Et “mystisk” bladmønster, en spiral med en lille divergensvinkel, undgår stadig beregningsmæssig forudsigelse, selvom Sugiyama mener, at de er tæt på at knække den bladrige kode.
”Vi synes ikke, vores undersøgelse er praktisk nyttig for samfundet, ” siger Sugiyama. ”Men vi håber, at det vil bidrage til vores forståelse af den symmetriske skønhed i naturen.”